【研究成果のポイント】
◆多くのカエル類では、オタマジャクシからカエルへの変態に伴って草食性から肉食性へと食性が変化し、その過程で胃と腸を区画化する幽門括約筋が形成されることが知られていたが、そのしくみは不明であった。
◆アフリカツメガエル(Xenopus laevis)における幽門形成過程の詳細な形態観察に加えて、in situ hybridization※1による遺伝子発現解析や、RNA-seq法による時期領域特異的かつ網羅的な遺伝子発現解析を行い、変態期に幽門括約筋形成が誘導されるしくみに迫った。
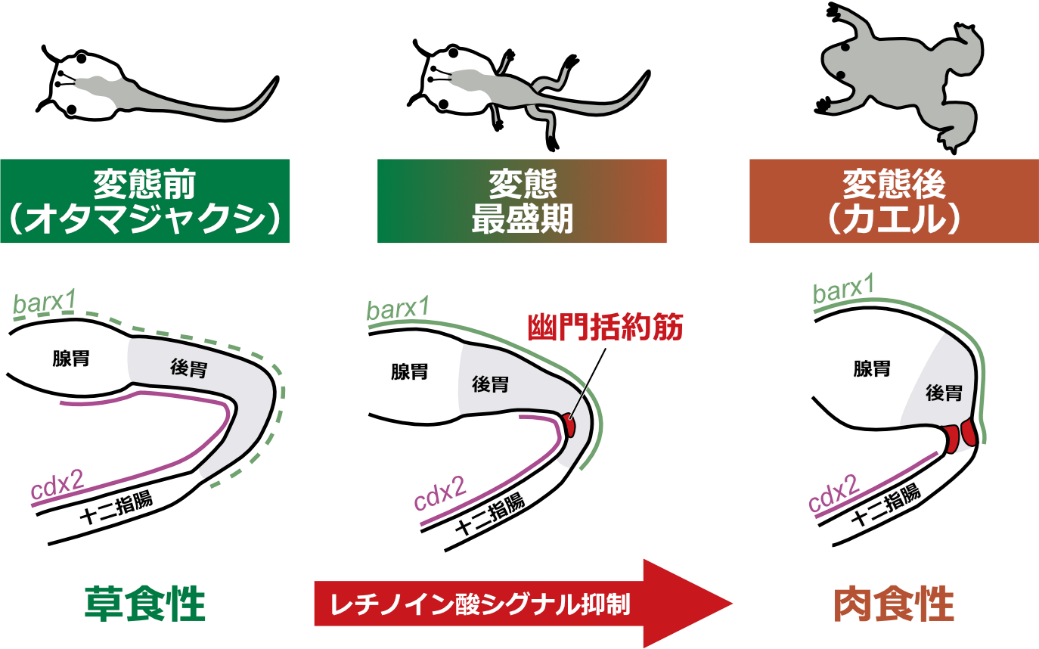
◆幽門括約筋形成の正確な時期と場所を特定したほか、変態期に胃の後方で発現変動する遺伝子を多数発見した。その結果、幼生期にはレチノイン酸※2シグナルが幽門括約筋形成を抑制し、変態期にその抑制が解除されることで幽門括約筋形成が始まる可能性が示された。
脊椎動物の消化管は食性に応じて多様な形態に進化しています。多くのカエル類では、幼生(オタマジャクシ)から成体(カエル)への変態に伴い、食性が草食性から肉食性へと変化します。変態期においては、肉食性に応じて腸の長さは4分の1に短縮し、腸管上皮は絨毛で覆われ、胃では蛋白質分解酵素のペプシンが分泌されるようになり、胃と腸を区画化する幽門括約筋が形成されます。これまで腸の変態の分子機構は盛んに研究されてきましたが、胃の変態の分子的研究はほとんどなく、特に、胚発生期※3や幼生期ではなく変態期に幽門括約筋形成が誘導されるしくみは未解析でした。
今回、東京大学大学院理学系研究科の名倉京大学院生(研究当時)と池田貴史大学院生(研究当時、現京都産業大学タンパク質動態研究所研究員)、平良眞規准教授(研究当時、現中央大学)らの研究グループは、アフリカツメガエル(Xenopus laevis)における幽門括約筋形成過程の詳細な形態観察を行い、幽門括約筋の形成位置や発達の過程を正確に記載しました。さらに、 前変態期と変態最盛期の網羅的遺伝子発現解析から、幽門括約筋形成に関与する可能性のある転写因子※4を多数同定したほか、レチノイン酸シグナルの制御が変態期特異的な幽門括約筋形成に重要である可能性を新たに見出しました(図1)。 本研究成果は、カエル類の変態とそれに伴う形態形成の新たなメカニズムを提唱するほか、食性に応じた消化管形態の多様化という進化生物学上の難問に答える手がかりを与えると期待されます。
本研究成果は2024年9月21日、米国発生生物学会の機関誌「Developmental Biology」にオンラインで掲載されました。
【用語解説】
※1 in situ hybridization
組織や胚において、目的の遺伝子が発現する(つまりRNAに転写される)場所を可視化する手法。RNAと相補的な配列をもつプローブを用意し、組織内のRNAと結合(ハイブリダイズ)させて検出する。
※2 レチノイン酸
ビタミンAを誘導体とし、レチナールデヒドロゲナーゼ(Aldh1a1)によって合成される小分子化合物。動物の発生において、前後軸の形成などさまざまな現象にかかわる重要なシグナル分子で、核内受容体の一種であるレチノイン酸受容体(RAR)に結合して下流の遺伝子発現を制御する。レチノイン酸はCyp26により分解され活性を失う。
※3 胚発生期
発生段階の中で、孵化までの時期か、あるいは摂餌開始前の時期。どちらの定義をとるかは種によって異なる。いずれにせよ形態形成がおこり多くの器官の原基がつくられる時期である。アフリカツメガエルでは、孵化した後も形態形成が進行するので、胚に口が開いて摂餌行動を始めるオタマジャクシとなるまでの時期とする。
※4 転写因子
DNAに直接結合し、遺伝子の転写を制御する機能をもつタンパク質。発生や細胞分化においては、異なる組織や細胞に異なる転写因子が発現し、それが結合する標的遺伝子の発現を促進または抑制することで細胞の分化の方向や最終分化を決定するなどの機能が知られる。
詳細は、大学ホームページの「プレスリリース」をご覧ください。